DNA hybridisation
We need to find genes that are causing problems so that we can alter them. This is done by DNA hybridisation: DNA probe is made by making a sequence of DNA is made that is complimentary to the gene and is fluorescent or radioactive (32p), the patients DNA is separated into two strands, the probe will bind to the gene and fluoresce or show up on a photographic plate to show where it is.
DNA sequencing
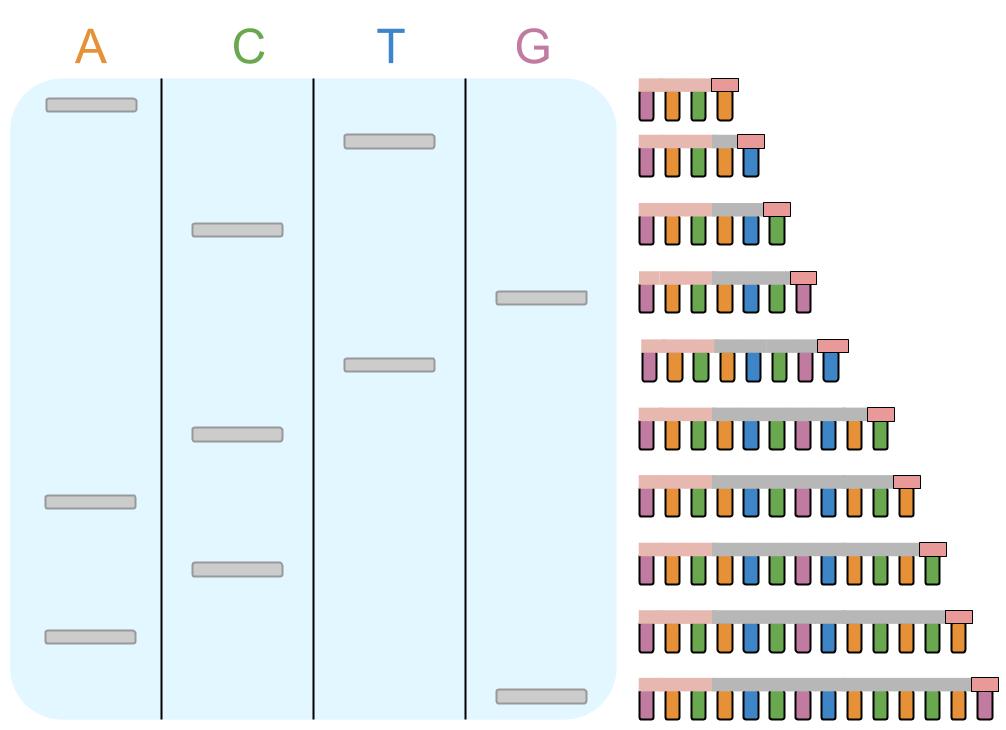
This is how to find out the order of bases of DNA:
- DNA is split into a single strand
- DNA polymerase is added
- Primers that are complimentary to the begining of the DNA are added (so that DNA polymerase has somewhere to start from)
- Free nucleotides are added
- One type terminator nucleotide is added (either C, T, A or G) that stops DNA polymerase when it is added to the chain; it will be radioactively marked (or more recently, fluoresce in a specific colour)
- DNA polymerase will make many complimentary strands of DNA, they may be stopped at some point in their making by the terminator nucleotide
- This results in many pieces of DNA of different lengths, each length corresponding to a place on the DNA where the base complimentary to the terminator nucleotide is
- This is done in four tubes, each one using a different terminator molecule
- So you will end up with a fragment that corresponds to each base on the sequence (however this cannot be seen yet)
- The mixture from each of the tubes is put on some agar gel, each in a separate column
- A voltage is applied to the gel, which makes the far end positive, which attracts the negative DNA strands (gel electrophoresis)
- Smaller molecule move more quickly and bigger molecules more slowly meaning that when the voltage is stopped the fragments are different distances through the agar base on size
- Photographic film is put over the top and detects where each strand is because of the radioactive marking (or more recently the agar is scanned with lasers by computers)
- This shows scientists the order of the complimentary bases so they can work out the sequence
Restriction mapping
DNA sequencing using the above method (sanger) can only be done using short fragments so DNA must be cut up using restriction endonucleases. Take for example this plasmid which has been cut up for sequencing:
Scientists cannot tell what order these fragments go in. In order to work it out they use the handy fact that restriction endonucleases only cut at specific sites.
To cut up this plasmid in the first place scientists would have used three restriction endonucleases, lets call them A, B and C. So, the whole plasmid is put in with just A and B and the lengths of the two resulting fragments recorded (using gel electrophoresis). This is then repeated but using just B and C, and then again using just A and C.
The whole plasmid is 10 kb (kilobases) long.
In the A and B trial there was one fragment of 2 and one of 8,
In the B and C trial one was 3 and the other was 7,
In the A and C trial there were two fragments of 5.
You can put all this information together to see that in between A and B must me 2 kb then in between B and C must be 3 kb and in between C and A must be 5 kb, because this adds up to 10. So now you can tell that the 2 kb fragment is first then the 3 kb fragment and then the 5kb fragment, and as the DNA for each of these fragments have been sequenced you can put it all together to know the sequence for the whole plasmid.
Sickle cell anaemia
This disease is caused by a substitution mutation on the gene for haemoglobin. It produces the amino acid valine instead of glutamic acid to make the protein haemoglobin s. Haemoglobin s has a sticky patch on it that makes it stick to other haemoglobin molecules making long, insoluble fibres (at low oxygen concentrations). This causes red blood cells to distort so they are sickle and inflexible meaning:
- They block capillaries
- They can't carry oxygen
- Malaria cannot exist on them
The mutated and non mutated genes be haemoglobin are co-dominant. If an individual has two genes for haemoglobin s they will be very ill and possibly die from it. If one gene for haemoglobin s and one for normal haemoglobin (a) an individual will be mostly ok, unless their oxygen concentration gets particularly low.
Being heterozygous for this trait is useful in places with malaria, because it is difficult for the parasite to survive in your blood and you are not severely ill.
DNA screening
This is looking at peoples genes to see if they are carrying a harmful mutation. This is done by adding a marked fragment of DNA that is complimentary to the mutated gene (a DNA probe) to a strand of DNA from the patient, if the patient has the mutated gene the probe will bind to it and show up because it is radioactive.
Genetic counselling
This is advice given to people who have found out they have a harmful, or potentially harmful mutation in their genotype due to genetic screening. For example a couple will be told what the likely-hood is of their offspring having certain problems, what the implications of these problems are and how they will know in the future if their offspring have the problem or not.
There are many applications of this concerning cancer:
- determining the best drugs to use to treat cancer (if the type of cancer is found out)
- determining the risk of relapse (if the amount of mutations are looked at)
- because two mutations are needed for a tumour to develop (oncogenes and inactive tumour suppressor cells) people may have one mutation, but not yet cancer, they can be told this to change their lifestyle in order to avoid the second mutation